Глава 10. Магний и фотосинтез
Ионы магния играют в поддержании жизни на Земле совершенно исключительную роль. Поток солнечной энергии, падавший на поверхность Земли в отдаленные геологические периоды, сначала не использовался примитивными формами живых существ. Положение, однако, постепенно изменялось, и некоторые из организмов приобрели особый аппарат, позволивший им направить этот практически неиссякаемый поток энергии в русло хемосинтеза, заставить свет участвовать в создании необходимых для клетки веществ.
Работа фотосинтетического аппарата позволяет клеткам, которые им вооружились, строить сложные органические соединения, прежде всего углеводы, из диоксида углерода СО2 и воды. В результате фотосинтеза в углеводах запасается энергия солнечного света:
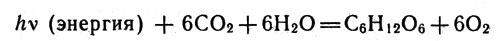
В результате фотосинтеза в углеводах запасается энергия солнечного света
Из уравнения видно, что кислород молекул воды освобождается, а водород идет на образование глюкозы, которая в дальнейшем превращается в крахмал. В процессах фотосинтеза, протекающих в клетках некоторых бактерий, источником водорода служит не вода, а другие вещества (например, сероводород).
Существенной частью биологического аппарата для использования энергии света является хлорофилл. Хлорофилл очень похож на гем: его молекула также содержит порфириновый цикл.
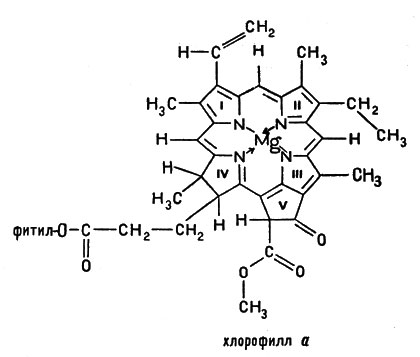
Хлорофилл очень похож на гем: его молекула также содержит порфириновый цикл
Отличие хлорофилла от гема заключается прежде всего в том, что хлорофилл - это комплексное органическое соединение магния, а не железа (как гем). Кроме того, в молекуле хлорофилла к порфириновому циклу присоединен еще и высокомолекулярный спирт - фитол. Известно несколько видов хлорофилла, но основной порфириновый каркас сохраняется во всех его видах. Есть и еще одно отличие гема от хлорофилла. Как видно из схемы, в молекуле хлорофилла, кроме типичных для порфирина четырех пиррольных колец, имеется дополнительный пятый цикл (V), содержащий атом водорода, карбонильную группу С=O и карбометоксильную группу 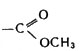 . В целом вся сложная макроциклическая система имеет ароматический характер; термин "ароматический" отнюдь не связан с каким-либо запахом - он указывает на сходство в состоянии электронов этого макроцикла и электронов ароматических углеводородов (а бензол и его гомологи действительно имеют легкий специфический запах). Часть электронов атомов углерода и азота порфиринового кольца - π-электроны - являются общими, так что внутри макроцикла существует своеобразный круговой ток (как и в бензоле).
. В целом вся сложная макроциклическая система имеет ароматический характер; термин "ароматический" отнюдь не связан с каким-либо запахом - он указывает на сходство в состоянии электронов этого макроцикла и электронов ароматических углеводородов (а бензол и его гомологи действительно имеют легкий специфический запах). Часть электронов атомов углерода и азота порфиринового кольца - π-электроны - являются общими, так что внутри макроцикла существует своеобразный круговой ток (как и в бензоле).
Атом водорода в цикле V активно вступает в различные реакции обмена.
Исследования свойств хлорофилла показали, что атом магния не находится строго в плоскости макроцикла, а выведен из этой плоскости, располагаясь над ней. Если хлорофилл растворить в жидкости, молекулы которой имеют полярное строение, то молекула растворителя присоединяется к магнию за счет своей пары электронов (такие молекулы называют электронодонорными)*. Если же среда, окружающая хлорофилл, неполярна, то молекулы хлорофилла соединяются друг с другом, причем роль электронодонорной группы выполняет группа >С=O цикла V (>С=O... Mg). В бензольном растворе существуют, например, двойные молекулы хлорофилла; в алифатических углеводородах, которые очень слабо присоединяются к магнию, образуются даже частицы, состоящие из десяти молекул хлорофилла. В воде хлорофилл практически нерастворим. Однако молекулы воды связывают молекулы хлорофилла вместе таким образом, что пара электронов атома кислорода воды взаимодействует с атомом магния одной молекулы, а атомы водорода воды образуют водородные связи с группами >С=O и -СООН другой (на схеме символ Сhl обозначает молекулу хлорофилла):
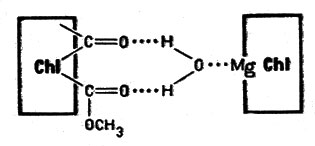
Пара электронов атома кислорода воды взаимодействует с атомом магния одной молекулы, а атомы водорода воды образуют водородные связи с группами >С=O и -СООН другой
* (Природа связи магния с лигандами в хлорофилле остается дискуссионной; по данным спектроскопических исследований, все четыре связи магния с азотом одинаковы и не имеют ионного характера.)
В результате возникают очень большие агрегаты частиц хлорофилла (коллоидные частицы). Следовательно, именно особенности строения макроцикла хлорофилла (наличие пятого цикла) и определяют его способность образовывать крупные скопления - агрегаты - тесно связанных частиц.
Свет действует на агрегаты хлорофилл - вода - хлорофилл, вызывает отделение электрона от одной из частиц комплекса, вода обеспечивает разделение зарядов.
В самой общей форме работу фотосинтетического механизма можно представить следующим образом. Молекула хлорофилла помещается между молекулами, способными присоединять и отдавать электроны. Свет, действуя на хлорофилл, переводит его электроны на более высокие энергетические уровни - свет поглощается хлорофиллом. (Возбужденное состояние обозначено на схеме звездочкой над символом хлорофилла Chl*). Затем возбужденный электрон выбрасывается молекулой хлорофилла и переходит к веществу, которое способно его принять, - так называемому акцептору электронов, и почти одновременно хлорофилл получает электроны от другого вещества - донора электронов:
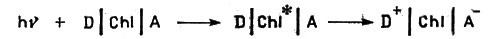
Работа фотосинтетического механизма
Здесь D - донор, А - акцептор электронов; hv - поглощаемый хлорофиллом квант света; Chl - хлорофилл.
Такие акты совершаются очень быстро: за одну секунду на солнечном свету происходит около 1500 вспышек. Между донором и акцептором поток электронов совершает круговой путь, расходуя свою энергию на разложение воды, образование АТФ и НАДФ*Н. Эта приближенная картина уточнялась в течение многих лет, но и поныне не все ее детали достаточно ясны.
В фотосинтетическом аппарате растений действуют две системы переносчиков и содержится хлорофилл двух видов, немного различающихся между собой. Свет с большей длиной волны (700 нм) действует на электроны хлорофилла типа а (система, обозначенная на рис. 22 цифрой I). Другая система (обозначенная цифрой II) содержит хлорофилл b и фикобилиновые пигменты. Она поглощает свет с меньшей длиной волны (680 нм). Именно в системе II разлагается вода и выделяется кислород. В каждой системе имеется по 200-250 молекул хлорофилла; только одна из них (в системе II), получая энергию фотона, передает электрон дальше - к веществу Q, остальной хлорофилл работает как светоулавливающее устройство.
Рассмотрим последовательность процессов фотосинтеза, представленных схемой на рисунке 22 и начнем с той части системы, которая обозначена цифрой II. В этой системе имеются группы молекул хлорофилла, на которые падает свет. В результате поглощения энергии света (квант hv) одна из молекул хлорофилла отдает богатый энергией (возбужденный) электрон веществу-переносчику, который обозначен на схеме буквой Q. Результатом потери электрона будет возникновение положительного заряда на молекуле хлорофилла (в системе II).
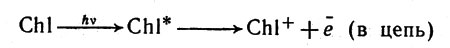
В результате поглощения энергии света (квант hv) одна из молекул хлорофилла отдает богатый энергией (возбужденный) электрон веществу-переносчику, который обозначен на схеме буквой Q
Для возвращения в исходное состояние молекула хлорофилла должна получить электрон. Установлено, что источником электронов является вода. Молекулы воды распадаются на ионы Н+ и ОН-. Ионы ОН- отдают свои электроны хлорофиллу, а сами превращаются в нейтральные группы - так называемые радикалы *ОН (каждый из которых содержит один неспаренный электрон). Эти радикалы взаимодействуют друг с другом с выделением кислорода. Считают, что в этих процессах важную роль играют ионы марганца.
Происходящие реакции можно схематически записать так:
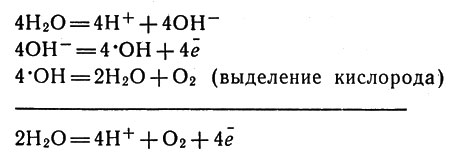
Последовательность процессов фотосинтеза
Но вернемся к электронам, текущим от молекулы вещества Q. Эти электроны, теряя свою энергию, двигаются затем по целому ряду переносчиков, которые мы не будем подробно описывать, и попадают в другую часть хлорофильной системы, обозначенной цифрой I. Важнейшим итогом их путешествия по этим переносчикам (до системы I) является образование АТФ, т. е. вещества, богатого энергией.
Заметим, что символом Р700 обозначена на схеме особая молекула хлорофилла, выбрасывающая возбужденные электроны. Среди переносчиков имеются соединения железа - железосерусодержащие и медьсодержащие белки (обозначенные на схеме FeS, Z), а также соединения хиноидной структуры (например, Q и пластохиноны).
В хлорофилльной системе I электроны снова возбуждаются светом и, следовательно, переходят на более высокий энергетический уровень. Их поток через переносчик Z (FeS-белок) и ферредоксин переходит к НАДФ+ и при участии ионов Н+ превращает кофермент в восстановленную форму:
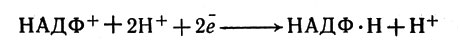
В хлорофилльной системе I электроны снова возбуждаются светом и, следовательно, переходят на более высокий энергетический уровень
В конце концов часть электронов может вернуться на путь, ведущий от переносчика Q к системе I, осуществив тем самым круговое движение электронов.
Итогом всей этой работы электронов, возбужденных светом, является, во-первых, образование АТФ, а во-вторых - получение НАДФ*Н за счет водорода воды. Оба соединения - и АТФ, и НАДФ*Н - необходимы для дальнейшей химической работы по созданию молекул углеводов. Назовем только основные этапы этих сложных реакций.
АТФ отдает энергию, способствуя образованию химически активного соединения углевода рибулозы - рибулозодифосфата (содержащего пять атомов углерода). Рибулозодифосфат, реагируя с диоксидом углерода, поступающего из внешней среды, превращается сначала в шестиуглеродное соединение, которое затем распадается на два трехуглеродных фрагмента - 3-фосфоглицерата:
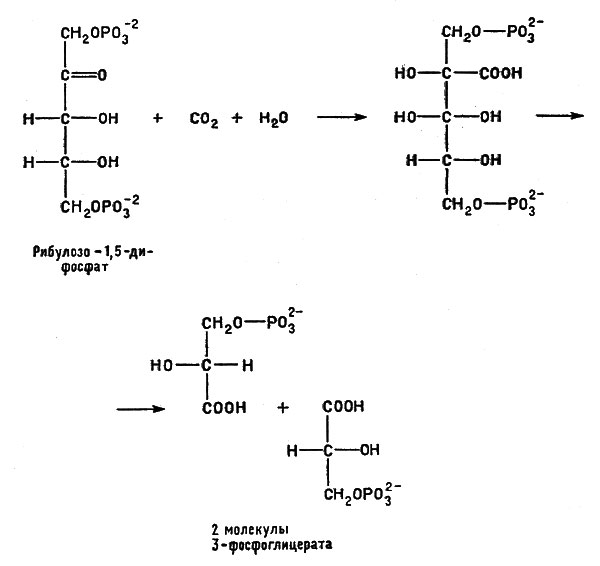
Рибулозодифосфат, реагируя с диоксидом углерода, поступающего из внешней среды, превращается сначала в шестиуглеродное соединение, которое затем распадается на два трехуглеродных фрагмента - 3-фосфоглицерата
Это соединение превращается с помощью НАДФ*Н в диоксиацетонфосфат, который является сырьем для синтеза фосфатов фруктозы, глюкозы и затем крахмала.
Синтез только 2 моль диоксиацетонфосфата, из которых образуется одна молекула глюкозы, требует, чтобы в фотохимической системе было произведено 12 моль НАДФ*Н и 18 моль АТФ. Следующие стадии синтеза глюкозы нуждаются в дополнительных количествах НАДФ*Н и АТФ.
Таким образом, образующиеся при фотосинтезе глюкоза и крахмал аккумулируют ту энергию, которую кванты света передали хлорофиллу на первых стадиях фотосинтеза.
В современной науке моделирование фотосинтеза - искусственное создание такой химической системы, которая могла бы, поглощая свет, доставлять нам углеводы, синтезируя их из воды и диоксида углерода, - является одной из самых увлекательных задач. Физики умеют превращать энергию света в электрическую энергию. Остается заставить поток электронов служить химическим целям с таким же совершенством, с каким это происходит в природе.
Для фотосинтеза нужен не только хлорофилл, но и особые мембранные структуры, содержащие белок и ряд ферментов в соответствующем пространственном расположении. Только до тех пор, пока все части этой сложной системы работают согласованно, идет процесс образования органических веществ, запасающих энергию света. В связи с этим познание фотосинтеза оказалось не только химической, но и физической проблемой и потребовало решения многих вопросов, в частности вопроса о передаче энергии возбуждения. Выяснилось, что структуры, необходимые для фотосинтеза, похожи на митохондрии: в зеленых растениях это хлоропласта, у микроорганизмов, многие виды которых способны к фотосинтезу (сине-зеленые, пурпурные бактерии и др.), все необходимые ферменты размещены в клеточной мембране, а мембрана впячивается внутрь клетки, образуя мезосомы.
Эти важные факты говорят о том, что структурная организация: возникновение надмолекулярных структур - митохондрий, хлоропластов - есть необходимое условие сопряжения реакций, доставляющих энергию, и реакций, поглощающих ее. Так в природе осуществляется переход от процессов на молекулярном уровне к процессам в макромолекулярных организациях - клетках и многоклеточных системах, причем и само существование таких систем поддерживается энергетическим сопряжением.
Все эти структуры образовались постепенно, и хлорофилл не сразу появился на Земле.
В осадочных породах, начиная с кембрийского, были обнаружены порфирины, и, как думают ученые, именно они являются остатками древних носителей хлорофилла. Некоторые ученые считают, что хлорофилл совершает свою работу в живых системах вот уже 550 млн. лет.
Хлорофилл и сам был рожден светом. Возможно, как считает М. Кальвин, солнечная радиация, действуя на воду и углекислый газ, привела сначала к появлению муравьиной и щавелевой кислот. Щавелевая кислота Н2С2О4 содержит два атома углерода. При облучении таких двухуглеродных молекул часто образуются четырехуглеродные молекулы; в частности, четыре атома углерода содержатся в кислоте С4Н6O4, называемой янтарной. С другой стороны, прямыми опытами Бахадура, А. Г. Пасынского с сотрудниками и других ученых доказано, что ультрафиолетовая радиация, действуя на растворы нитратов и формальдегида, в которых содержатся соли железа, вызывает появление в растворах аминокислот; аминокислоты могут возникнуть и при действии электрических разрядов на смесь азота, углекислого газа и паров воды. Из янтарной кислоты и аминокислоты глицина, NH2CH2COOH, по-видимому, и образовались порфирины.
Они и были (по А. А. Красновскому) первичными аппаратами для использования энергии света. У наиболее древних видов бактерий - автотрофов - обнаружено наличие свободных порфиринов. Бактерии действовали в лишенной кислорода восстановительной атмосфере, которая была характерна для ранних периодов истории Земли, и содержали восстановленную форму порфирина. Однако свободные порфирины, в силу особенностей их спектра поглощения, не могут обеспечить достаточно полного использования видимой части солнечного излучения. Постепенно химическая эволюция усовершенствовала аппарат и привела к образованию хлорофилла; внедрение магния в структуру порфирина вызвало повышение активности, и, кроме того, магний укрепил связи хлорофилла с белком*.
* (Некоторые ученые (Гранин) считают, что сходство гема и хлорофилла обусловлено тем, что оба вещества при биосинтезе сначала образуются одинаковым путем, и лишь на более поздних стадиях их синтез идет по-разному; различают "железную" и "магниевую" ветви: образование с одной стороны гема, с другой - хлорофилла.)
Фотосинтез в той его форме, которая приобрела особенно большое значение на Земле, совершается в зеленых частях растений и в водорослях. Трудно представить колоссальные масштабы деятельности зеленых водорослей, плавающих в морях и океанах. За один год они выделяют в атмосферу Земли 3,6*10 т кислорода. Это составляет около 90% всего кислорода, поступающего в атмосферу из океана и с поверхности листьев наземных растений. Следовательно, именно водоросли являются основным поставщиком кислорода. Именно они создают ту часть атмосферы, которая необходима для нашего существования. Наша жизнь неразрывно связана с "дыханием океана" и непрерывной деятельностью микроскопических носителей хлорофилла. К. А. Тимирязев в книге "Солнце, жизнь и хлорофилл" изложил результаты своих фундаментальных исследований в области фотосинтеза и указал, что фотосинтез - это процесс, от которого зависят все проявления жизни на нашей планете.
Развитие фотосинтетических аппаратов знаменовало собой начало совершенно нового периода в эволюции форм жизни на Земле. Появились новые виды живых существ, резко изменились условия питания, состав атмосферы - началось обогащение ее кислородом. Синтез органических веществ в растениях и водорослях обеспечил пищей гетеротрофные* организмы; из остатков растений под влиянием химических и биологических факторов начали образовываться массы ископаемых углей. Накопления таких отложений, как нефть и сланцы, - это тоже результат фотосинтетической деятельности.
* (Способные жить за счет питания готовыми органическими соединениями; отличаются от автотрофов - способных первично синтезировать необходимые им вещества.)
Пожалуй, невозможно найти другой биохимический аппарат, который мог бы с таким совершенством использовать энергию излучения для химических целей, как это делает хлорофилл. Хлорофилл действительно является звеном между энергией Солнца и жизнью на Земле; этим определяется исключительная роль ионов магния в развитии форм жизни.
Лишь очень небольшая часть энергии, падающей на лист, используется им для целей фотосинтеза, эта часть не превышает 1%. И тем не менее общая продукция фотосинтеза на Земле колоссально велика. Биохимики называют фотосинтез самым крупным из химических процессов на Земле. Действительно, продукция фотосинтеза за год (1011 т органического углерода) в 100 раз превышает ежегодную мировую продукцию угольной и нефтяной промышленности; энергия, которую накапливают фотосинтетические машины в организмах, в 10000 раз превышает энергию воды, используемую в гидростанциях всего мира, и в 100 раз - энергию сгорания угля.
Эти числа, конечно, являются ориентировочными; но тем не менее их порядок дает вполне ясное представление о масштабах биологического процесса, который называется фотосинтезом.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://iznedr.ru/ 'Из недр Земли'